本文是一篇临床医学论文,我们的研究加深了对调控离皮质投射神经元亚型规范的激活、抑制转录网络的时间和分子性质的理解,并为FOXG1综合征感觉运动功能障碍的发病机制提供重要线索。
第一章文献综述
1.大脑新皮质投射神经元的亚型多样性及其功能
哺乳动物的大脑新皮质是神经系统中最为复杂精细的结构,含有百余种不同类型的神经元,这些神经元定位于特定的皮质层,形成特异的联系和投射,构成复杂的神经环路,在神经发育过程中,参与调控机体感觉运动、语言认知、社交情感等生命活动。
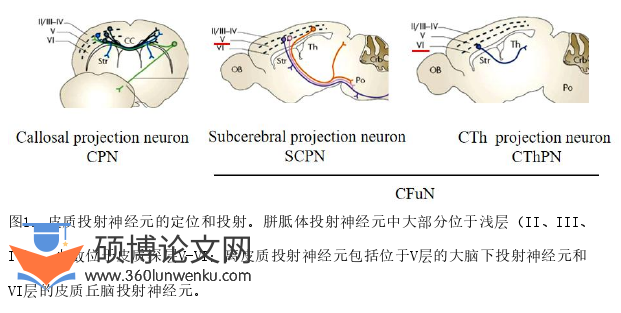
大脑新皮质由不同类型神经元排列组成典型的六层结构。大脑皮层的兴奋性投射神经元也是按照投射特性分层排列。浅层主要是胼胝体投射神经元,参与双侧半球信息整合。深层主要是离皮质投射神经元(corticofugal neurons,CFuNs),是大脑皮质的主要信息输出系统。CFuNs根据定位和投射分为2类,定位于第VI层,投射到丘脑的皮质丘脑投射神经元(corticothalamicalprojection neurons,CThPNs)和V层投射到脑干和脊髓的大脑下投射神经元(subcerebral projection neurons,SCPNs)[1,2](图1)。
临床医学论文怎么写
..........................
2.大脑新皮质投射神经元产生的时序性和皮层定位
在神经发育过程中,大脑新皮质的神经发生始于胚胎期10.5(E10.5),随着时间进程将有序产生不同类型的神经元并定位形成六个独特的皮质层,每一层的神经元都具有其独特的形态特征、电生理特性、轴突投射类型和基因表达模式[23,24]。早期产生的神经元会向外迁移离开增殖区,形成前板(preplate,PP),之后产生的神经元则迁移入前板,将其分隔成底板(subplate,SP)和边缘层(marginal zone,MZ),而在底板和边缘层中间则聚集形成皮质板(corticalplate,CP)。随着神经发生,新生神经元将从内到外依次产生并迁移排列,定位形成典型的六层结构(图2)。早产生的神经元最先定位于新皮质深层(V、VI层),后产生的神经元则迁移越过已产生的深层神经元,定位于皮质浅层(II、III、IV层)[25-28]。这种时间依赖性模式决定了皮质神经元的层定位和亚型特性,是皮质投射神经元多样化的基础。在神经发生过程中,E11.5是底板神经元产生的高峰期,E12.5开始产生定位于VI层的皮质丘脑投射神经元,E13.5产生V层的脑下投射神经元,E14.5之后主要产生II-IV层的皮质投射神经元[29,30]。离开细胞周期的有丝分裂后神经元开始启动层特异性基因的表达,VI层皮质丘脑投射神经元CThPNs高表达Tbr1,V层大脑下投射神经元SCPNs特异性表达Bcl11b(Ctip2),浅层胼胝体投射神经元CPNs高表达Satb2。在这些特异性基因的调控下皮质细胞最终特化为不同类型的神经元,并建立起精确的神经环路连接,提示有丝分裂后神经元亚型特化调控机制是个十分复杂且逐步精细的过程,在神经元命运获得中发挥着重要作用。
............................
第二章材料与方法
1.材料
1.1实验动物
本研究所用小鼠:ICR小鼠,Foxg1fl/fl基因小鼠,NEX-Cre小鼠和CAG-CreER小鼠。其中NEX-Cre小鼠信息见文献所述[130,131],CAG-CreER购自与Jax lab。所有小鼠繁殖饲养均在东南大学实验动物中心开展,严格按照《实验动物管理条例》执行。实验不考虑小鼠性别差异,且所有的实验小鼠都在ICR背景上进行解析。
1.2主要仪器或设备
水平电泳槽(0603,日本)恒温水浴锅(HH-S2,江苏金坛医疗仪器厂)高压蒸汽灭菌锅(LDZX-30FBS,上海申安医疗器械厂)电热恒温干燥箱(DHG-9146,上海精宏实验设备有限公司)恒温电热板(DB-IVA,江苏金坛医疗仪器厂)超净工作台(VS-1300L-U,苏州安泰空气技术有限公司)激光共聚焦显微镜(Olympus,FV1000)荧光显微镜(DP71,Olympus)荧光显微镜(EVOS FL auto,Life)紫外分光光度计(Nano drop2000,Thermo scientific)PH仪(PHS-3BW,上海般特仪器制造有限公司)脱色摇床(TY-80S,南京大学普阳科学仪器研究所)原位杂交炉(HM-4000,USA)漩涡振荡器(Genie2,USA)全自动雪花制冰机(IMS-85,常熟市学科电器有限公司)冰箱(DW-FL270,中科美菱低温科技有限责任公司)医用冷冻冷藏冰箱(YCD-EL259,中科美菱低温科技有限责任公司)冰冻切片机(CM3050S,Leica)
......................
2.实验方法
2.1小鼠交配策略
将Foxg1fl/fl小鼠与NEX-Cre小鼠交配即可获得NEX-Cre;Foxg1cKO的条件性敲除小鼠。Foxg1fl/fl小鼠与CAG-CreER小鼠交配,需在胚胎期E14.5注射他莫昔芬诱导,最终获得CAG-CreER;Foxg1cKO的条件性敲除小鼠。(交配策略见下图)

临床医学论文怎么写
2.2交配和鼠龄确定
一般取2个月龄的小鼠进行交配,保证其性成熟且状态良好。在下午15:00至17:00将雌雄小鼠进行合笼,并于次日上午10:00前检测雌鼠是否形成阴道栓,至少连续检测3天,应将检测到阴道栓的日期标记为E0.5天,将小鼠出生的日期标记为P0天。
.........................
第三章实验结果.....................34
1.离皮质投射神经元特化过程中,SCPNs和CThPNs亚型特异性基因的表达模式变化................................34
2.Foxg1在离皮质投射神经元中高表达............................35
第四章总结和讨论..........................53
1.Foxg1在CFuNs亚型特化中的机制研究.................53
2.FOXG1综合征与CFuNs功能缺陷................................55
第三章实验结果
1.离皮质投射神经元特化过程中,SCPNs和CThPNs亚型特异性基因的表达模式变化
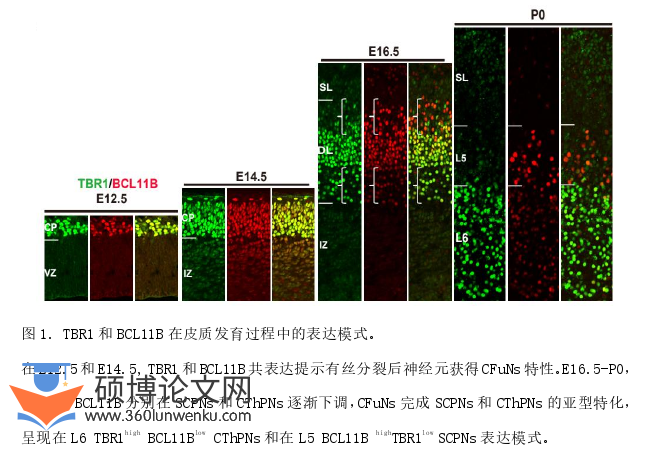
哺乳动物的新皮质是典型的六层结构,在皮质发育过程中,神经元依次产生并准确迁移定位到特定皮质层,分化为特定亚型。研究报道,神经发生早期(E14.5之前)主要产生的是定位于深层的神经元,之后分化为定位于L5、L6的SCPNs和CThPNs。为了进一步确定离皮质投射神经元亚型特化的时间进程,以及在此过程中亚型特异性基因的表达模式变化,我们首先检测了E12.5-P0的CThPNs亚型特异基因Tbr1和SCPNs亚型特异Bcl11b的表达模式变化。结果表明:在E12.5,TBR1和BCL11B共表达于皮质板的有丝分裂后神经元,呈现出TBR1highBCL11Bhigh的表达模式。从E14.5开始,TBR1highBCL11Bhigh神经元明显增加,但在皮质板上部部分神经元中TBR1的表达逐渐减少。在E16.5,TBR1和BCL11B表达分化为三层模式,皮质板上部为TBR1weakBCL11Bhigh神经元,提示这些神经元将获得SCPNs亚型特性;在皮质板下部为TBR1highBCL11Blow神经元,提示这些神经元将特化为CThPNs;中间仍有部分TBR1highBCL11Bhigh的共标神经元,提示为正在特化中的CFuNs。在P0时期,皮质深层呈现出典型的第5层(Layer 5,L5)BCL11BhighTBR1lowSCPNs和第6层(Layer 6,L6)BCL11BlowTBR1highCThPNs的分布模式(图1)。这些结果提示在E14.5之前产生的有丝分裂后神经元共表达TBR1和BCL11B,获得CFuNs的特性,在E14.5之后,TBR1和BCL11B分别在预定为SCPNs和CThPNs的神经元中被下调,从而完成CFuNs向SCPNs和CThPNs的特化。
临床医学论文参考
..............................
第四章总结和讨论
1.Foxg1在CFuNs亚型特化中的机制研究
在本研究中,我们首先确认了层特异性分子Bcl11b和Tbr1在皮质深层神经元中的表达情况,早期有丝分裂后神经元TBR1highBCL11Bhigh表达模式提示这些神经元获得CFuNs的亚型特性,在P0时期皮质板呈现出第五层BCL11BhighTBR1lowSCPNs和第六层BCL11BlowTBR1highCThPNs的表达模式,表明Bcl11b和Tbr1的时空表达的精确调控决定着CFuNs向SCPNs和CThPNs的精准特化。Foxg1作为端脑发育过程中表达最早的基因,在E8.5天就在皮质神经上皮中表达。在神经元特化过程中,Foxg1在早期CFuNs,以及后期SCPNs和CThPNs的持续高表达,提示Foxg1可能参与Bcl11b和Tbr1的时空特异性表达,并调控CFuNs向CThPNs和SCPNs亚型特化的进程。
前期报道表明在有丝分裂后神经元中激活Bcl11b和Tbr1的表达能调控CFuNs的命运特化。为了在E14.5之后完成TBR1highBCL11BlowCThPNs特化,必须降低BCL11B在CThPNs中的表达。在CThPNs中,FEZF2/BCL11B的异位高表达可以逆转其特化进展,最终将其转换为SCPNs命运。我们的研究发现FOXG1作为负调控因子,在E14.5之后可以直接抑制Bcl11b转录,控制其在CThPNs中的表达水平,促进CThPNs的亚型特化。同样的,为了在E14.5之后完成TBR1lowBCL11BhighSCPNs特化,在开启SCPNs命运时需要降低TBR1表达水平。FOXG1作为Tbr1转录的阻遏物,在E14.5之后通过直接抑制Tbr1的转录,下调SCPNs中TBR1水平,促进SCPNs的亚型特化。
参考文献(略)