第 1 章 文献综述
1.1 PED 的临床症状、致病机理及病理变化
PEDV 可导致所有年龄阶段的猪只感染发病,1 周龄以内的仔猪感染后主要表现为水样腹泻、呕吐、脱水,死亡率高达 100%。断奶仔猪感染后表现为呕吐和厌食,死亡率约为 1%-3%,育肥猪或成年种猪感染后主要表现为厌食和水样腹泻,持续 3-5 天左右,很少会造成死亡。PEDV 感染猪只后,主要定植在小肠粘膜上皮细胞内,并在其中大量复制,PEDV感染 24 小时内,通过破坏小肠上皮细胞,引起猪只肠绒毛萎缩和脱落(Jung et al2014),而在感染猪的结肠内未能发现组织病理学变化。PEDV 主要感染消化系统,病变主要集中在小肠段,剖检可见小肠肠壁变薄、透明,内充满淡黄色液体,并可造成小肠黏膜萎缩、脱落,严重时出现肠壁充血,其他组织器官无肉眼可见明显病理变化。本病无明显的季节性,一年四季均可发生,但寒冷的冬春季节更易发生。PEDV主要通过粪-口途径传播,健康猪只直接或间接接触被病毒污染的水源、饲料、饲料原料、车辆或者未消毒干净的栏舍都可引起新生仔猪发病(Pasick et al 2014),也有证据表明,PEDV 能够通过气溶胶传播(Alonso et al 2014)。有学者报道,在感染母猪的乳汁中也能检测到 PEDV(Li et al 2012; Sun et al 2012b),仔猪可以通过吃母乳感染病毒,这为切断病毒的传播增加了难度。
..........
1.2. 单克隆抗体研究进展及应用
1.2.1 单克隆抗体概述
单克隆抗体(monoclonal antibody,McAb)于 1975 年首次问世,在疾病的预防、诊断和治疗方面发挥越来越重要的作用。目前,单克隆抗体多为鼠源性,而鼠源性单抗存在使宿主产生免疫反应、在宿主体内半衰期较短等临床应用的局限性。随着分子生物学技术的发展,实现了单克隆抗体由鼠源向人源的转变,嵌合型单克隆抗体、人源化单克隆抗体、小分子抗体和全人源单克隆抗体应运而生。这些技术在临床诊断和疾病治疗方面得到广泛应用。本文主要综述单克隆抗体在猪传染性疾病中的应用及进展。通过制备PRRSV GP3蛋白的单抗,利用ELISA方法鉴定GP3的抗原表位,实现了对 GP3 抗原结构的分析(Zhou et al 2006)。Wang 等人利用噬菌体筛选技术,在单克隆抗体和多克隆抗体的基础上,绘制了非洲猪瘟病毒主要衣壳蛋白线性和构象表位的图谱(Wang and Yu 2009)。Meng 等于 2010 年成功获得 11 株针对 PCV2Rep-N 的单抗,成功鉴定到 L(39)FDYFIVG(46)和 K(99)EGNLLIE(106)两个线性表位,其中 L(39)FDYFIVG(46)和 K(99)在 PCV2 中高度保守,而PCV1 在此处存在 V(41)C(42)氨基酸替换。这为 PCV2 诊断试剂盒的研制和PCV Rep 蛋白功能的解析奠定了基础(Meng et al 2010)。单克隆抗体在疾病诊断中既可以用于抗体检测,又可以用于抗原检测。主要将单克隆抗体与ELISA或免疫层析试纸条相结合。Huang等人研制出了以PCV2单克隆抗体为基础的阻断 ELISA,该方法操作简单、特异性强、敏感度高,既可以用于临床 PCV2 分子流行病学监测,又可以用于评价 PCV2 中和抗体水平(Huang et al2011)。在基于 FMDV 非结构蛋白 NSP 3B 单克隆抗体和兔源多克隆抗体的基础上,Fu 等研制出了固相阻断 ELISA(SPB-ELISA),用来区分 FMDV 疫苗免疫和自然感染(Fu et al 2014)。Morioka 等利用单克隆抗体建立的夹心 ELISA,用于临床 FMDV O 型抗体监测和诊断(Morioka et al 2009)。以杆状病毒截短表达的TGEV-S 重组蛋白和 3 株特异性的单克隆抗体为基础,成功开发了一种区分 TGEV与其他猪源呼吸道冠状病毒的抗原捕获 ELISA(Lopez et al 2009)。Li 等利用针对JEV E 蛋白的两个单抗 2A2 和 4D1,建立了一种快速检测临床 JEV 感染的免疫层析试纸条(Li et al 2010)。
..........
第 2 章 PEDV 单克隆抗体的制备及应用
2.1 研究目的及意义
由 PEDV 变异毒株导致的 PED 大规模暴发,给世界养猪业造成了巨大经济损失,同时也引起了各国学者的广泛关注。PEDV 快速检测方法的建立及致病机理的研究在 PED 防控中显得尤为重要。本研究制备了针对 PEDV S1D 蛋白的单克隆抗体,可以与 PEDV 病毒粒子发生特异性反应,在用于临床 PEDV 检测的同时,也为PEDV 基础性研究提供材料。
......
2.2 试验材料
CO2培养箱(SANYO)、台式离心机(Eppendorf centrifuge,5415R)、倒置显微镜(Olympus)、倒置荧光显微镜(Olympus IX70)、Millipore 超纯水机(18Mn.cm,Milli-Q,Mimpore)、分光光度计(Spectrum SHANGHAI 765Pc)、超净工作台(AIRTECH)、高压破碎仪、SDS-PAGE 电泳仪(Bio-Rad)、WesternBlotting 转膜仪(Bio-Bad)、凝胶成像系统(Bio-Rad)、冰冻切片机(Leica)、石蜡切片机(Leica)、脱水机(Leica)、石蜡包埋机(Leica)等。HAT 盐、HT 盐、50% PEG、弗氏完全佐剂和弗氏不完全佐剂、淋巴细胞分离液、二甲基亚砜(DMSO),均购自 Sigma 公司,RPMI1640 培养基购自 Hyclone公司,胎牛血清(FBS)和青霉素-链霉素溶液购自 GIBCO 公司,羊抗鼠 IgG-HRP抗体购自 ABclonal 公司,FITC 标记羊抗鼠 IgG 抗体购自 SouthernBiotech 公司。小鼠抗体亚类鉴定试剂盒(Pierce Rapid ELISAMouse mAb Isotyping Kit)购自Thermo scientific 公司。rProtein G Beads 购自常州天地人和生物科技有限公司。
........
第 3 章 PEDV 强弱毒株感染 Vero 细胞的蛋白质组学分析..........43
3.1 研究目的及意义........... 43
3.2 试验材料............. 43
3.2.1 细胞、毒株 ........ 43
3.2.2 主要试剂与试剂盒 ...... 43
3.2.3 主要培养基及溶液的配制 .... 44
3.2.4 主要缓冲液及相关试剂的配制 ...... 44
3.2.5 主要实验器材 .... 44
3.2.6 分子生物学信息软件 ............ 45
3.3 试验方法............. 45
3.4 结果与分析......... 55
3.5 讨论 ...........75
3.6 小结 ...........76
第 4 章 PEDV 感染 Vero 细胞诱导细胞自噬的机制研究.....77
4.1 研究目的及意义 ...........77
4.2 试验材料 .............77
4.3 试验方法 .............78
4.4 结果与分析......... 83
4.5 讨论........... 93
4.6 小结........... 95
第 5 章 全文总结.........96
第 4 章 PEDV 感染 Vero 细胞诱导细胞自噬的机制研究
4.1 研究目的及意义
PEDV 感染 Vero 细胞蛋白质组学分析发现,ATG5、微管结合蛋白 1B 等自噬相关蛋白在PEDV感染过程中显著上调,同时自噬相关通路mTOR显著下调,这提示细胞自噬可能与 PEDV 在 Vero 细胞上的增殖有关。研究表明,细胞自噬在多种病毒的复制过程中发挥重要作用,而在PEDV复制中的作用研究还未见报道。探究细胞自噬在 Vero 细胞增殖过程中的具体作用,可以为解析 PEDV 致病机理以及新型抗病毒策略的开发提供新的视角。抗 LC3、Beclin l、ATG5、p62、β-actin 抗体以及实验室所用辣根过氧化物(HRP)标记二抗均为 ABclonal 公司产品;抗 P65、p-P65 抗体为 Cell SignalingTechnology(CST)公司产品;抗 PEDV S1D 单克隆抗体为本实验室自制(参见第2 章); 抗 HA 标签抗体为 Medical & Biological Laboratories(MBL)公司产品;Alexa Fluor 488 标记(绿光)以及 Alexa Fluor 594 标记(红光)驴抗鼠 IgG 购自武汉安特捷生物技术有限公司;DAPI(4',6'-diamidino-2-PhenyIinciole)为碧云天生物技术研究所产品。
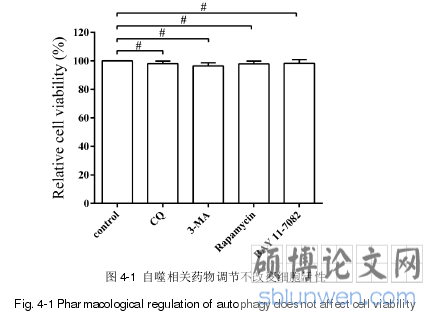
.......
总结
猪流行性腹泻病毒变异毒株的出现,给世界养猪业造成了巨大经济损失,其致病机理的研究在新型抗病毒策略的制定中具有非常重要的意义。本文围绕围绕猪流行性腹泻病毒开展了系列研究,主要结论如下:
1. 成功获得 3 株可以特异性识别 PEDV 病毒粒子的针对 PEDV-S1D 蛋白的单克隆抗体。在此基础上,建立了PEDV快速诊断方法,用于临床PEDV肠道样品检测,且该方法敏感性显著高于 RT-PCR 检测。
2. 获得 PEDV 强弱毒株感染 Vero 细胞的蛋白表达谱,进一步分析发现,差异蛋白主要聚焦于蛋白质合成、免疫调控、细胞组装、信号转导和凋亡等。
3. 较弱毒株而言,PEDV 强毒株显著上调 NF-κB 通路,诱发更强烈的炎症反应。此外,PEDV 强毒株通过下调 mTOR 及其下游靶分子 4EBP1 和 p70S6K,以及eIF2(真核起始因子)通路活性来抑制细胞蛋白质的合成。
4. 本研究首次报道了 PEDV 感染 Vero 细胞可以诱导完全细胞自噬进程,进而促进PEDV 的复制。此外,自噬可能参与了 PEDV 介导的炎症反应,与 NF-κB 信号通路诱导的炎症反应存在正向调控关系。
..........
参考文献(略)