1 前言
1.1 PRRSV 总论
猪繁殖与呼吸综合征(PRRS),是由猪病原猪繁殖与呼吸综合征病毒(PRRSV)引起的一种诱发母猪孕后期流产、产死胎、产木乃伊胎(周庆雨,2010),幼仔患呼吸系统疾病和大量死亡的传染病(Bautista et al., 1993;Wensvoort, 1993;Voice and Kolb,1994)。因发病猪群中部分猪耳部呈紫色,故又称猪蓝耳病(周建国,2013)。二十世纪九十年代世卫组织在专家讨论会上决定国际统一采用 PRRS 这一命名。该病最早发生于二十世纪八十年代的美国(杨汉春,2016),后相继在北美一些国家被发现,而后又出现于欧洲等多个国家。随着时间的推移,大洋洲中以澳大利亚为代表的等多个国家境内也相继发生该传染病(Nam et al., 2009)。1991 年,我国台湾省出现该传染病(徐彦召,2012),3 年后,日本爆发猪繁殖与呼吸综合征。1996 年,我国大陆出现该传染病,由郭宝清团队于爆发流产的猪胎儿中分离并鉴定(郭宝清等,1996)。之后,该传染病在我国境内迅速蔓延开来,在我国内陆、沿海、东北、江南等省份均分离到了 PRRSV 病毒株(宁宜宝等,2006),自此,在我国以 JXA1 毒株为代表的高致病性毒株在猪群中造成了大规模的发病率和死亡率(Tong et al., 2007;Li et al., 2007;Tian et al., 2007)。因该传染病最早爆发于北美洲和欧洲,故而分类上又常以北美型和欧洲型分类(Adams et al.,2014;Van Doorsselaere et al., 2012),此两大类病毒的基因序列约有 50%的同源性(Fanget al., 2007;Forsberg, 2005)。近些年来,美国一直流传着一种严重危害生殖系统的疾病,被称为“急性”或称“非典型” 猪繁殖与呼吸综合征(李毅,2012),由此说明,出现了一种毒力更加强、传染力更加大的新型 PRRSV 病毒株,美国一直流传的新型传染病也表明了 PRRSV 一直在不断的变异(刘晓东,2019)。
该病毒株的遗传变异多样性导致了我们对于 PRRSV 的防疫防控对策更加地难以制定(刘翠权,2011)。PRRSV 在猪群间的传播方式多种多样。分类为水平和垂直传播两种方式。猪群间水平传播的方式又被分为直接和间接的两种接触传播方式(顾小雪,2015)。有关 PRRSV 流行病学研究表明猪群中发生的以直接接触方式发生传播的案例多以猪繁殖与呼吸综合征猪的分泌物被健康猪接触为主。氯仿和醚类等脂溶剂能够使病毒囊膜破坏以致病毒难以复制从而丧失活性(Benfield et al., 1992;Bloemraad et al.,1994)。 除去我们所知的短距离间接接触传播方式外,PRRSV 也可通过空气流动远程传播。
..........................
1.2 IL-4 研究进展
1.2.1 IL-4 起源
白细胞介素 4,最早被发现于 1982 年,当时的研究人员发现培养 T 细胞的上清液中含有一类能促进 B 淋巴细胞增殖的因子(Paul, 1991;Postlethwaite and Seyer, 1991)。起初将其命名为 B 细胞生长因子-1(BCGF-1)、也有研究人员将其命名为 B 细胞刺激因子-1(BSF-1)、T 细胞生长因子-2(TCGF-2),1986 年,人工将其基因成功克隆(任力强,1990),此后,国际统称为白细胞介素 4(interleukin 4,IL-4)。
1.2.2 IL-4 的来源与功能
IL-4 一直是人类设计有效的小分子抑制剂的理想目标(陈德高,2019)。这类细胞因子主要由嗜碱性粒细胞,嗜酸性粒细胞和肥大细胞以及 Th2 淋巴细胞产生(袁芳,2014)。除此以外,自然杀伤(naturalkiller,NK)T 细胞、人类胃上皮细胞同样产生 IL-4(涂云霞,2020),在小鼠中,主要由 T-helper-2 细胞产生 IL-4。因其翻译后再修饰的不规则模式,尤其是糖基化,其分子量变化范围在 12~20KDa(邢应如,2015)。这类细胞因子与IL-13 类细胞因子共同享有绝大部分细胞表面受体,序列的同源性,细胞内信号转导和以及有限的功能作用等(林亚迪,2018)。大量研究表明,IL-4 参与特定的细胞增殖,多种基因的表达,并且阻止各种细胞类型(如淋巴细胞、巨噬细胞、上皮细胞、成纤维细胞以及内皮细胞)的凋亡(于潇,2019)。在 IL-4 蛋白与其他一些细胞因子(IL-13,IL-5和 IL-10 等)蛋白的协同作用下,促进 Naive T-helper 细胞分化和转化为 T-helper-2 细胞。IL-4 会强烈停止 T-helper-1 细胞分化,并且下调其产生 IFN(王欣欣,2020)。 IL-4 的另一个主要功能是调节并转化免疫球蛋白的类别,特别是调节人类 B 淋巴细胞中 IgE 和IgG4 的表达以及调节小鼠 B 淋巴细胞中和 IgE1 和 IgG1 表达。在 IL-4、IL-4 受体(IL-4R)、STAT-6 敲除的小鼠体内,IgE 的产生量减少至少 100 倍。一致的认为 IL-4 能够增强病原特异性诱导的体液免疫应答(Li et al., 2015),IL-4 通过作为 B 淋巴细胞生长的促有丝分裂原,并在造血组织中具有多重作用(王奔放,2018)。通过与脂多糖的结合使用,可增加 IL-4 受体、CD 23、MHC II 类分子的表达,并且增强 B 淋巴细胞上 ThY-1 的表达。IL-4 还可以延长培养的 T 和 B 淋巴细胞的寿命(程侠卫,2013),但不会单独充当静止淋巴细胞的生长因子(马占川,2020)。它还可以通过因子依赖性骨髓血统来避免 IL-4表达细胞的凋亡。
...............................
2 材料和方法
2.1 材料
2.1.1 细胞和病毒株
mPAM-CD163(本实验室构建的含有 CD163 受体的猪肺泡巨噬细胞系)、PBMCs(分选自首次感染 JXA1-R 株 21 dpi、42 dpi、84 dpi 仔猪外周血)、CD4+T 细胞(分选自首次感染仔猪外周血 PBMCs)、HP-PRRSV JXA1 毒株(中国动物疫病预防控制中心惠赠)。
2.1.2 试验仔猪
试验仔猪,5 头 4-6 周龄健康长白二元杂交断奶仔猪,购自泰安某 PRRSV 双阴性种猪场,饲养于泰安某小型猪场。
2.1.3 试验小鼠
SPF 级 C57BL/6L 小黑鼠购自济南朋悦试验动物繁育有限公司,7-8 周龄、雌性、体重 17-20g,饲养于山东农业大学 SPF 试验鼠房,试验鼠房恒温 25℃,相对湿度40%-70%,昼夜明暗交替时间为 12h/12h,投放无菌鼠粮,灭菌水。
2.1.4 主要试剂和耗材

医学论文参考
2.2 仔猪首次感染 PRRSV 相关试验方法
2.2.1 首次感染仔猪方案
仔猪的饲养管理、免疫程序、血液采集时间
5 头 4-6 周龄健康长白二元杂交断奶 PRRSV 双阴性仔猪,购买当日,运输至某地某小型猪场,运输当天,禁食,少量给予饮水。次日,采集血液样本送至某高校试验室。运用 PCR 技术检测 PRRSV 抗原阴性,IDEXX ELISA 试剂盒检测结果,抗体同为阴性。饲养于独立猪舍,参考猪的饲养管理标准:保育阶段,维持猪舍内温度在 22-25℃,相对湿度在 65%-75%,应保证每头仔猪每天 1-2L 的饮水量,1000-2000g 的采食量。育肥阶段,维持猪舍内温度在 19-22℃,相对湿度在 65%-75%。应保证每头仔猪每天 3-5L的饮水量,1800-2500g 的采食量。于独立猪舍中正常饲养管理 3d,逐日观察仔猪精神状态,采食饮水情况,记录体温指数等。各项指标均正常后,接种高致病性猪繁殖与呼吸综合征病毒活疫苗(JXA1-R 株),首次感染日期为 0 dpi。21 dpi、42 dpi、84 dpi,无菌采集仔猪外周血,试验具体操作如下:
一名试验人员用套猪器套住试验仔猪上颚,另一名试验人员迅速用 75%酒精棉球擦拭试验仔猪颈部以下嵌窝处,然后用 12 号无菌兽用采血针管采集外周血,轻轻推入已经用 75%酒精棉球擦拭过顶部的 5mL 枸橼酸钠无菌抗凝管中,边往无菌抗凝管中推血液边轻轻旋转混匀,最后轻轻把无菌抗凝管颠倒混匀,颠倒混匀 20 次左右。无菌采集的抗凝血,务必于 2h 内送至原代细胞间进行 PBMCs 的分离,时间越久,分离效果越差。
医学论文怎么写
.............................
3 结果与分析.................................34
3.1 PRRSV 首次感染仔猪后 PBMCs 中 CD4+ T 细胞比例、淋巴细胞转化率、PBMCs 体外培养后猪 IL-4 表达水平............................ 34
3.1.1 首次感染仔猪外周血 PBMCs 中 CD4+ T 细胞比例....................34
3.1.2 首次感染仔猪外周血中淋巴细胞转化率............................35
4 讨论.......................52
5 结论.........................................60
4 讨论
猪繁殖与呼吸综合征(PRRS)俗称“蓝耳病”,引起该症状的病原体为猪繁殖与呼吸综合征病毒(PRRSV)。在发生于猪群的呼吸道疾病中,该疾病一直处于主要地位(Chenet al., 2017)。PRRSV 基因组全长 15.1kb,属于单股正链 RNA 病毒,该病毒基因组编码 8 个结构蛋白和 14 个非结构蛋白(Li et al., 2007)。同时,PRRSV 能使所有年龄段的猪群感染,主要引起发高热、嗜睡、呼吸困难、食欲不振等临床症状(Jiang et al., 2000),其中怀孕母猪妊娠孕后期多发生早产、流产、产死胎、木乃伊胎及弱仔等临床症状(Burkard et al., 2018)。
自 1995 年在我国分离第一株 PRRSV 毒株和 2006 年在我国分离第一株 HP-PRRSV毒株以来,该疾病在我国迅速传播,给我国的猪肉生产商上带来了巨大的经济损失(Tianet al., 2007)。至 2012 年,NADC30 毒株在我国境内流行变得更加普遍(Li et al., 2016),严重增加了对该疾病的预防和控制难度(Wang et al., 2017),给我国造成了更加巨大的经济损失。一方面 PRRSV 进入宿主后,导致宿主的持续性感染,继而导致 PRRSV 在猪群中大规模蔓延(朱迎春, 2020);另一方面 PRRSV 不断出现新的突变体和不同毒株间极易发生毒株重组(方莹, 2012)。客观上来讲,已有研究对 PRRSV 来源、进化、与宿主相互作用关系的诠释是不充分的。尽管市场上已有疫苗(减毒活疫苗和灭活疫苗)对 PPRSV 进行预防,但这此两种疫苗存在不同的缺点。减毒活疫苗具有良好的保护性,但 PRRSV 的频繁变异导致活疫苗有极其强的毒株特异性,同时也存在病毒的基因组和毒力返强的风险;灭活疫苗的安全性极好,但是对细胞免疫的诱导不够充分,同时也不能够提供足够的保护力(胡悦,2016)。近几十年对 PRRSV 的研究进展显示对 PRRSV发病机制的研究是不充分的,因此进一步研究 PRRSV 的发病机制是极其有意义的。
.............................
5 结论
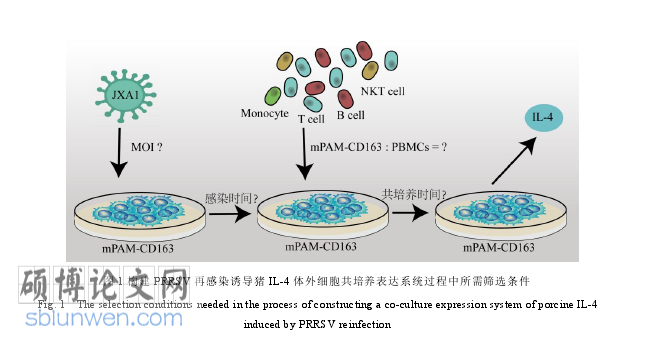
5.1 建立起 PRRSV 再感染诱导猪 IL-4 体外表达的细胞共培养系统,系统条件为MOI=0.01,病毒感染 1h,mPAM-CD163:PBMCs=1:5,细胞共培养 18h,该系统下体外再感染组中出现 CD4+T 细胞与 mPAM-CD163 细胞相互结合现象。
5.2 初步建立了 PRRSV 诱导 IL-4 表达的小鼠模型,试验 14d 首次免疫组小鼠脾脏中淋巴小结数量增多,形状增大,均匀分布。
参考文献(略)