本文是一篇农业论文,本论文初步对PITG_02860和Cu/ZnSOD(PITG_15626)在致病疫霉中生物学功能进行了研究,发现Cu/ZnSOD(PITG_15626)亚细胞定位发现其位于菌丝和孢子囊的细胞核和细胞质中,与致病疫霉的生长发育、内吸性农药胁迫、氧化胁迫及致病疫霉的侵染能力相关。
第1章前言
1.1马铃薯概述
马铃薯(Solanum tuberosum L.)是茄科茄属一年生草本植物,富含大量的蛋白质、淀粉和多种维生素等对人体有益的营养成分,在作为主食、预防疾病等方面不断发挥出重要的应用价值并具有非常高的经济利用价值。我国马铃薯种植面积是世界第一,但我国的马铃薯的单亩产量却依然低于全球种植的平均水平。在种植过程中不可控的环境影响和难以预防的病害都会引起马铃薯产量下降。在19世纪50年代的爱尔兰,就因为马铃薯晚疫病的严重爆发,导致马铃薯的产量损失严重,进而引发了饥荒,人们饱受饥饿,流离失所。晚疫病爆发,是毁灭性的灾难,它的爆发可以使马铃薯种植面积大范围死亡,严重危害经济发展。因此,有效防治马铃薯晚疫病对于马铃薯的经济产业提高是必须的。
1.1.1马铃薯起源与传播
马铃薯别名洋芋、荷兰薯、番仔薯、爪哇薯、地蛋、山药等。马铃薯的传播对人类影响深远,拓展人们的生存和发展空间,促进了社会的发展历程[1]。关于马铃薯的来源,目前的观点是马铃薯由秘鲁南部的印第安人种植,后从秘鲁传播到南美大陆安第斯山区[2]。十六世纪三十年代马铃薯被第一次引入西班牙[3],西班牙的马铃薯很快就蔓延到了欧洲和亚洲的许多地方。传入欧洲后,土豆适应了较短的生长季节,并在漫长的日子里形成块茎。十九世纪中叶,由于港口的开通,许多国家的移民、传教士、商人、探险家和游客涌入,马铃薯的输入和引入渠道日益增多[4],马铃薯的种类也越来越多。马铃薯引入中国已有300多年的历史[5],中国是世界马铃薯主产区之一。谢从华[6]等推测马铃薯最早传入中国的时间在可能在1423年(郑和第六次航海时),在清代时传播开来。祝菊澧等[7]认为云南省并不是马铃薯传入中国的起源地,其主要观点为:(1)马铃薯在1615年由印度传入滇西,由滇西开始传播到云南省各处。(2)1788年后经四川传入楚雄,开始在滇西及滇中传播。(3)1843年经贵州传入昭通在滇东北开始传播。
........................
1.2马铃薯晚疫病
1.2.1马铃薯晩疫病及病源概述
自然界中,马铃薯的生长发育主要受到各类自然因素和生物因素的影响。首要的是马铃薯种植环境的湿度、温度、光照、土壤等。马铃薯在全球范围内都有栽培,根据当地的气候和地理环境,其种植技术也各有差异。一般为以块茎为原料,进行无性生殖。马铃薯也容易受到生物因素的侵染,菌物类病害主要是晚疫病、疮痂病、早疫病等;细菌性病害主要有环腐病和青枯病等;病毒病主要有花叶病、卷叶病、类病毒病和纺锤形块状病毒病等,主要害虫有草地贪夜蛾、块茎蛾、线虫、蓟马、地老虎等[23]。由卵菌纲(Oomycetes)致病疫霉菌(Phytophthora infestans)引起的晚疫病危害马铃薯的生长发育[24-25],一般年份产量损失在50%左右,严重时绝收,马铃薯晚疫病严重影响马铃薯的产量,严重威胁人类生存及全球粮食安全[26-28]。由晚疫病造成的马铃薯种植业的巨大损失最早可追溯到19世纪40年代,爱尔兰地区的晚疫病大爆发,马铃薯产量损失过半引发了大饥荒,大量爱尔兰人因饥饿死亡并导致100万人的逃荒他国[24]。此后,晚疫病在世界各马铃薯主产区频繁发生,其危害呈上升趋势。因此,控制晚疫病、提高马铃薯产量和品质仍然是全球农业生产面临的巨大挑战。
致病疫霉菌丝的结构特点及其发病规律特点与真菌极其相似,导致前期将其误归属为真菌界。经过进一步的研究发现,其归属于卵菌门(Qomxcota),卵菌纲(Oomycetes),霜霉目(Peronosporales),腐霉科(Pythiaceae),疫霉属(Phytophthora)[29],二倍体异宗配合,基因组大小240 Mb,也发现三倍体形式存在[30]。
......................
第2章致病疫霉原生质体制备及转化系统研究
2.1实验材料试剂
2.1.1实验材料
致病疫霉菌株88069(A1交配型亲本),由华中农业大学田振东教授惠赠,放置于18℃恒温培养箱内保存生长。pTOR-mRFP由南京农业大学王源超教授惠赠。
2.1.2试验仪器和试剂
仪器:移液枪、高温高压灭菌锅、PCR仪、基因扩增仪、电泳仪、恒温水浴锅、恒温培养箱、台式显微镜、超净工作台、离心机、激光共聚焦显微镜等。
试剂:DNA快速提取试剂盒、一次性注射器、细菌过滤器、葡萄糖、蔗糖、黑麦培养基、PM豌豆培养基、PA豌豆培养基等。

农业论文怎么写
......................
2.2实验方法
2.2.1原生质体的制备条件优化
(1)致病疫霉的培养和繁殖:用手术刀切致病疫霉88069菌块于90 mm黑麦-V8固体培养基培养11天。
(2)在固体培养基上新鲜生长培养的88069致病疫霉,沿菌落边缘打孔10mm的菌丝块,挑10块菌丝块置于装有10 mL的离心澄清的豌豆液体培养基(90mm的培养皿)中,于18℃培养2-3d,过滤收集菌丝体,用灭菌的0.8 mol/L甘露醇冲洗2次后投入35 mL 0.8 mol/L甘露醇中于26℃40 rpm摇洗10 min,再用灭菌的纱布过滤,收集的菌丝体加入细胞壁裂解酶液(用细菌过滤器过滤)20 mL,于26℃、40 rpm条件下裂解30 min。消化后的细胞悬浮液经灭菌的两层Miracloth(Calbiochem)过滤,即得到原生质体。通过原生质体的量确定最佳培养时间,取最佳培养时间菌丝,18℃,酶解30分钟为基础进一步探究裂解酶和纤维素酶在不同浓度比下的原生质体制备率;确定裂解酶和纤维素酶的浓度后,在此基础上进一步优化酶解时间。最后,以上述最佳条件为基础,本研究通过温度梯度实验,得到P.infestans原生质体最佳制备条件。过滤后的细胞悬浮液用封口膜封好管口于18℃2000 rpm离心3 min,轻轻倒出上清,不用全倒完,否则原生质体会损失很多。加入35 mL W5溶液,轻轻晃动两下沉淀就能被悬浮起来。重悬原生质体后细胞悬浮液用封口膜封好管口18℃2000 rpm离心4 min,轻轻倒出上清。加入5mL W5溶液,用封口膜封好管口冰浴30 min。在冰浴的同时,吸出原生质体在显微镜下观察计数。
...............................
第3章Cu/ZnSOD(PITG_15626)的功能鉴定....................................20
3.1实验材料.............................20
3.1.1实验材料......................................20
3.1.2试验所需仪器和试剂.......................20
第4章PITG_02860表型功能分析...........................35
4.1实验材料....................35
4.1.1实验材料...............................35
4.1.2试验所需仪器和试剂.......................35
第5章总结与展望.............................45
第4章PITG_02860表型功能分析
4.1实验材料
4.1.1实验材料
致病疫霉P.infestans菌株88069(A1交配型亲本),127P、126P、124P和121P(有性生殖诱导F2代A1型),88133(A2交配型亲本),110P、112P、117P和118P(有性生殖诱导F2代A2型),88069由华中农业大学田振东教授惠赠,88133由中国农业大学国立耘教授惠赠,其余菌株由云南师范大学马铃薯科学研究院前期诱导和放置于18℃恒温培养箱内保存生长。马铃薯B7材料由实验室提供种植于校内马铃薯研究院基地。pTOR-mRFP由南京农业大学教授惠赠。
4.1.2试验所需仪器和试剂
仪器:离心机、移液枪、高温高压灭菌锅、PCR仪、核酸浓度测定仪、恒温摇床、荧光定量仪、基因扩增仪、电泳仪、恒温水浴锅、恒温培养箱、游标卡尺、台式显微镜、激光共聚焦显微镜等。
试剂:LB液体培养基、反转录试剂、高保真酶(TaK aR a公司)、TB GreenPremix Ex TaqⅡ(TaK aR a公司)、琼脂糖凝胶回收试剂盒(DP219-03)、细菌过滤器、黑麦培养基、质粒小提试剂盒(TianGen)、PM豌豆培养基、PA豌豆培养基。
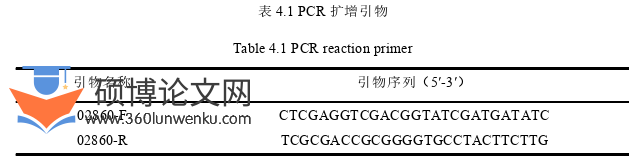
农业论文参考
............................
第5章总结与展望
本论文初步对PITG_02860和Cu/ZnSOD(PITG_15626)在致病疫霉中生物学功能进行了研究,发现Cu/ZnSOD(PITG_15626)亚细胞定位发现其位于菌丝和孢子囊的细胞核和细胞质中,与致病疫霉的生长发育、内吸性农药胁迫、氧化胁迫及致病疫霉的侵染能力相关。发现PITG_02860与致病疫霉生长、产孢量、卵孢子形态和毒性有关,与内吸性农药胁迫无显著相关性。
同时本研究也探索了致病疫霉PEG转化方法,菌丝用碗豆汁培养3天,采用复合酶(Lysing enzyme,10 mg/mL;cellulase,5 mg/mL)于26℃裂解45min,原生质体制备率最高为31.5±2.36个/μL;原生质体再生率可达3.34%±0.20%。以载体pTOR-mRFP质粒转化,通过PCR验证和荧光观察确认转化成功。
结合进一步探索优化PEG转化方法(针对质粒种类和质粒浓度进行改良),提高原生质体再生率,可针对Cu/ZnSOD(PITG_15626)和PITG_02860构建靶向突变和过表达相关载体,利用优化的PEG方法对其生物学功能开展进一步研究。此外,可在植物中挖掘两个重要基因的互作蛋白,深入了解Cu/ZnSOD(PITG_15626)和PITG_02860在致病疫霉侵染时的互作机制。
参考文献(略)