本文是一篇农业论文,本研究将含有预测靶向结合位点的假基因RPS4XP1和FOXO1 3'UTR野生型与突变型基因片段连接至pSiCHECK2基因报告载体,并分别与miR-223共转染至Hela细胞中进行双荧光素酶活性检测。结果表明,miR-223与RPS4XP1和FOXO1 3'UTR分别靶向结合。
1 前言
1.1 非编码RNA的研究进展
基因表达是生命各个方面所需的中心过程,根据经典的中心法则,基因组DNA首先转录成RNA,然后RNA翻译成蛋白质,最终来行使生物学功能。RNA通常被认为是连接DNA与蛋白质的遗传信息桥梁,是遗传信息传达的唯一信使。大量的转录组研究表明,大约三分之二的哺乳动物基因组被积极转录,而真核生物的基因组中大约只有1.9%转录产物编码蛋白质,超过90%转录产物由非编码RNA(no-coding,ncRNA)构成,不具备编码蛋白质的能力[1-4],根据长度、形状、位置的不同,ncRNA可被分为不同的类别。如近年热门研究的微小RNA(miRNA)、小干扰RNA(siRNA)、长链非编码RNA(lncRNA)、环状RNA(circRNA)以及PIWI相互作用RNA(piRNA)[5-11],同时还有研究较为冷门的假基因(pseudogene)[12-14]。然而ncRNA表达量水平低,序列保守性差,缺乏完整的开放性阅读框架(Open Reading Frame,ORF),所以常被认为是“转录噪音”。1969年,Davidson与Britten提出的真核细胞各类RNA可通过Watson-Crick互补配对原则来调控基因功能[15],众多研究也表明RNA不仅仅只起到转录过程中遗传信息的传递媒介功能,随着近年来高通量、深度转录组测序技术的快速发展,越来越多的ncRNA被发现在转录、转录后和翻译水平上调控编码基因的表达水平,间接参与调控各种生物学功能[4,16]。
1.1.1 LncRNA概述
LncRNA通常是指一类几乎不编码蛋白、转录本长度大于200个核苷酸(nucleotide,nt)的RNA。如图1-1所示,lncRNA按照其在基因组中相对于蛋白质编码基因的位置可大致分为正义、反义、基因内和基因间lncRNA(lincRNA)等几种类型。LncRNA可以从增强子转录而来(elncRNA),它们大多是双向转录,一些lncRNA也可通过mRNA或其他lncRNA的内含子反剪接产生环状RNA(circRNA)[17]。LncRNA与mRNA一样,由聚合酶II转录,并被5’端加帽、聚腺苷酸化以及剪接[18]。大多数lncRNA通过聚腺苷酸化稳定[19],而非聚腺苷酸化的lncRNA可以通过二级结构稳定,例如其3’末端的三螺旋结构[20]。但平均而言,它们含有的外显子数量少于mRNA,并且它们在不同组织中的表达水平较低,所有哺乳动物lncRNA几乎没有共同的结构、功能或机制特征。尽管RNA序列保守性较差,但lncRNA在共生基因组区域的物种中经常被鉴定出来,在跨物种间发挥相似的功能[21]。
................................
1.2 FOXO1基因的研究进展
1.2.1 FOX家族简介
叉头蛋白家族(Forkhead box,FOX)是一类功能庞大、高度保守的转录调控因子家族,不仅参与调节基因转录与DNA损伤应答反应,还参与调控细胞分化、增殖、凋亡、自噬以及胚胎的生长发育等众多的生命进程。随着对其研究的深入,从酵母杆菌到哺乳动物的FOX蛋白家族,研究者们根据其特定的DNA结合结构域和其他功能序列将FOX家族分为由FOXA至FOXS的19个亚家族,超过100个家族成员[75]。其中叉头框蛋白O(Forkhead box protein O,FOXO)是FOX家族中的一个亚家族,是胰岛素及胰岛素样生长因子受体下游的进化保守途径的关键参与者。哺乳动物的FOXO家族包括FOXO1,FOXO3,FOXO4和FOXO6,它们在结构、功能及调控机制方面具有高度的相似性[76]。
1.2.2 FOXO1的研究进展
1990年,Anderson等人首次在人类肺泡横纹肌肉瘤的染色体易位中鉴定得到FOXO1,并表明了其存在重要调控作用[77]。如图1-7所示,FOXO1在结构上包含4个功能域:(1)forkhead功能域(DBD):该区域为DNA结合域,对于核靶向至关重要;(2)核定位信号域(NLS):该区域显示了CBP/P245的乙酰化位点(K248、K262和K300)、AKT的磷酸化位点(T24、S256和S319)和14-3-3的结合位点(T24和S256);(3)核输出信号域(NES):调节核易位;(4)转录激活区域(TAD):反式激活域[78]。其中FOXO1的二级结构包含4个α螺旋(H1、H2、H4和H3),3个β折叠(S1、S2、S3)和2个翼环(W1和W2),有利于FOXO1 蛋白与靶标DNA保守识别序列(TTGTTTAC)相互作用,促进转录[79,80]。
............................
2 材料与方法
2.1 实验材料
2.1.1 主要仪器
本实验所用到的主要仪器如表2-1所示。

农业论文怎么写
.............................
2.2 实验方法
2.2.1 原代奶牛乳腺上皮细胞的体外分离、培养与鉴定
2.2.1.1 原代DCMECs的体外分离与培养
原代DCMECs培养大多采用酶消化法、组织块培养法、机械法与酶消化法的组合培养法、其它的各种方法与组合培养法等。本实验采用如下酶消化法体外分离培养DCMECs。
(1) 取样前准备:将取原代所需广口瓶、烧杯、锥形瓶、手术刀、剪刀、镊子、止血钳、铜网、离心管等器材分别高压湿灭与高温干灭进行灭菌处理;DMEM/F12原代培养液(5×双抗、2.5 μg/mL两性霉素B、20%胎牛血清),D-Hank’s原代缓冲液(10×双抗、2.5 μg/mL两性霉素B);培养瓶至少提前8 h铺鼠尾胶原溶液(120 μg/mL),置于37℃、5% CO2细胞培养箱中。
(2) 在超净工作台中将取回的奶牛乳腺实体组织用10×D-Hank’s清洗2次,洗掉血渍和奶渍,再用剪刀剪成多个小块,用氟康唑溶液浸泡清洗2次,最后再用10×D-Hank’s清洗数次,尽可能洗去奶渍,并浸泡其中,留后续备用。
(3) 用剪刀剔除乳腺组织表面的结缔组织、血管、乳导管以及脂肪,将剩余富含DCMECs的实质组织剪成约1 mm³的小块,再用10×D-Hank’s清洗数次至澄清。
(4) 将剪碎的组织放入锥形瓶中,加入纯培养液和I型胶原酶,封口,避光,置于37℃、140 r/min摇床消化3 h。
(5) 在超净工作台中把消化好的细胞混合液用100目铜网过滤2次,将滤液置于15 mL离心管中,1000 r/min离心10 min。
(6) 弃上清,将细胞沉淀用10×D-Hank’s吹匀,1000 r/min离心10 min重复两次。
(7) 弃上清,将细胞沉淀用DMEM/F12原代培养液吹匀,分至已铺胶原的细胞培养瓶中,置于37℃、5% CO2细胞培养箱中。
(8) 培养3~5天观察细胞贴壁状况,并进行纯化处理。
..........................
3 结果与分析...................................... 36
3.1 DCMECs 原代培养及鉴定 ..................................... 36
3.2 miR-223 靶基因的筛选与鉴定 ....................... 37
4 讨论 ........................................ 66
4.1 假基因RPS4XP1 的编码性与保守性 ....................... 66
4.2 假基因 RPS4XP1、miR-223 与 FOXO1 互作的调控网络 ..... 67
5 结论 ........................... 75
4 讨论
4.1 假基因RPS4XP1的编码性与保守性
近年来,随着生物信息学和高通量测序技术的迅猛发展,越来越多的ncRNA被挖掘鉴定。其中假基因由编码蛋白基因演化而来,是与同源基因序列相似的一段DNA,目前已鉴定出数以万计的假基因,其中假基因转录产物通常被认作是lncRNA,其生物学功能也被越来越多研究所报道[104-107]。LncRNA通常低于mRNA的表达水平,mRNA通常在物种之间具有高度保守性,然而lncRNA由于其产生方式,通常序列保守性差,但共生基因组产生的lncRNA常被鉴定出来,在跨物种间发挥相似功能。目前我们通常认为假基因是由正常基因(真基因)通过反座或复制两种方式形成与真基因序列相似的一段DNA。假基因被认为因受选择压力而累积突变产生的结果,使我们看不到其原有的基因,成为一些无规则的序列,被封存或获得新功能。依据同源性搜索结合共现性分析表明,人和猕猴之间存在保守的转录假基因,而在不同物种之间保守性强于侧翼序列[108]。这同样表明假基因在探索基因多样化的产生、物种亲缘关系、基因进化等方面有着重要意义。本研究通过运用NONCODE与LncPep数据库分析发现,假基因RPS4XP1在牛和人类物种中存在序列保守性,具有很高的可靠性。
通常我们认为lncRNA不可编码蛋白质,然而最近也有研究表明lncRNA可以编码蛋白并发挥功能,如St-Germain等人表明,假基因Pfh8-ps可以在体内产生PHF8-PS蛋白[46]。Dubois等人表明假基因UBBP4可编码UbKEKS[109]。随着蛋白质基因组学的核糖体图谱和质谱分析证实,假基因衍生的长非编码RNA可能是编码蛋白的重要来源,但是蛋白质水平上的假基因功能很少被考虑[110]。因此在研究一个新lncRNA时,除了检测全场序列,探究其是否编码蛋白质也是必要的。如Li等人通过通过转录组测序发现了一种新的lncRNA,并将其命名为lncRNA ADNCR,通过将其序列构建于pET-28a原核过表达载体,体外翻译系统并未发现蛋白质产物,证明为非编码RNA,通过研究发现该lncRNA ADNCR可通过靶向miR-204调控牛脂肪细胞分化[30]。本研究将假基因RPS4XP1构建于pET-28a原核表达载体上,与阳性对照组pET-28a-RFP及阴性对照组相比,经诱导剂诱导表达后,体外翻译系统发现假基因RPS4XP1并未翻译出蛋白质产物,因此假基因RPS4XP1为非编码RNA。
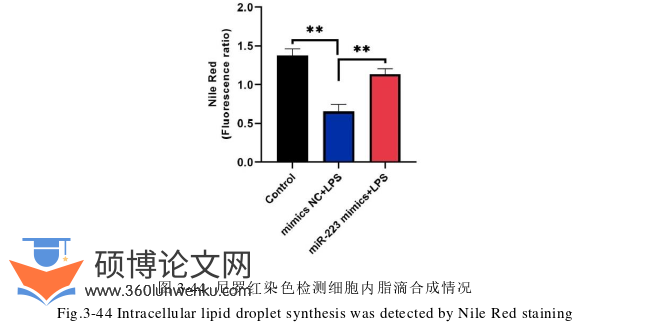
农业论文参考
................................
5 结论
(1) 假基因RPS4XP1不可翻译蛋白质,为非编码RNA,并且在人和牛物种之间存在序列保守性。
(2) 假基因RPS4XP1、miR-223与FOXO1之间存在ceRNA调控网络。
(3) 假基因RPS4XP1可充当miR-223的分子海绵,通过激活AKT/FOXO1、AKT/mTOR信号通路,进而调控DCMECs增殖、凋亡、乳脂合成。
(4) LPS刺激DCMECs可促进FOXO1蛋白和miR-223基因的表达;miR-223在DCMECs中具有抵抗LPS诱导的炎症反应的作用,恢复对乳脂合成的抑制作用。
(5) 假基因RPS4XP1可能通过靶向miR-223调控铁死亡相关基因NOX3、CA9和ALOX15表达,可能促进DCMECs铁死亡发生。
参考文献(略)